Le muscle est un tissu formé de fibres douées de contraction permettant de produire un mouvement. Les muscles participent à diverses fonctions essentielles à la vie de l’organisme (respiration, digestion…) et à son adaptation au milieu environnant (locomotion, thermorégulation…).
La macroscopie des muscles et leur type de contraction a permis de distinguer deux grands groupes de muscles : les muscles rouges, striés volontaires ou muscles de la vie de relation et les muscles blancs, lisses involontaires ou muscles de la vie végétative.
Les caractéristiques fonctionnelles et anatomiques des différents muscles permettent de les subdiviser en quatre grandes catégories.
- Muscle Lisse
- Muscle strié cardiaque
- Muscle strié squelettique
- Un exemple de muscle strié à insertion conjonctive : le sphincter de l’urètre
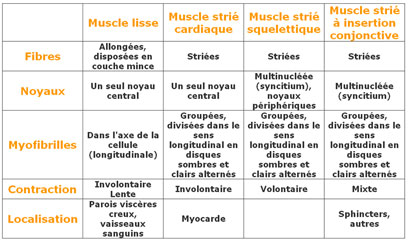
Tableau des différents types de muscles
Muscle lisse
Description macro et microscopique
Les muscles lisses sont présents dans la paroi de nombreux organes (tous les vaisseaux sanguins sauf les plus petits, intestins, utérus…). Ils forment des couches denses qui tapissent la paroi interne des vaisseaux et des organes creux et ne montrent pas de stries transversales. Ils sont constitués de cellules fusiformes mononucléées de taille variable (20 à 200 µm) dont le noyau est en position centrale, les fibres musculaires lisses. Ces cellules sont soit isolées dans le tissu conjonctif, soit regroupées en tunique musculaire (vaisseaux, tube digestif) ou en muscles (muscle érecteur du poil). Généralement, les faisceaux des fibres lisses des tuniques musculaires sont organisés en deux couches superposées : une couche circulaire et une couche longitudinale. L’orientation de ces couches est définie par l’orientation des fibres musculaires lisses par rapport à l’axe de l’organe.
Métabolisme et vascularisation
Le muscle lisse en général dépend plus du métabolisme anaérobie. Ainsi les muscles lisses des parois artérielles sont avasculaires. Ce n’est pas le cas des muscles lisses du tube digestif.
Innervation
Les muscles lisses sont sous le contrôle du système nerveux neurovégétatif (ou système autonome) qui ne répond pas au contrôle de la volonté. Les fibres nerveuses du système autonome présentent, dans le muscle lisse, des varicosités axonales qui se présentent sous la forme de renflements en forme de bulbe. Ces varicosités libèrent les neuromédiateurs nécessaires à la stimulation des fibres musculaires lisses dans une fente relativement large : les jonctions diffuses. Cette stimulation induit ainsi la contraction des fibres musculaires lisses.
Contraction
Une contraction rythmique caractérise les muscles lisses unitaires ou muscles viscéraux qui ne sont pas adaptées à la réalisation de mouvements fins. Les fibres lisses de ces muscles sont couplées électriquement entre elles par l’intermédiaire de « jonctions à trous » (« gap junctions »). Ces muscles viscéraux fonctionnent donc comme des syncytiums même s’il n’existe pas de ponts protoplasmiques entre ces cellules : on parle donc de syncytium fonctionnel. Ils montrent spontanément des contractions irrégulières et continues indépendantes de l’innervation. Cet état est appelé tonus. Ces muscles sont impliqués dans le péristaltisme.
Une contraction graduée caractérise les muscles lisses multiunitaires qui se retrouvent par exemple dans l’iris de l’œil. Les fibres musculaires lisses qui constituent ces muscles sont indépendantes les unes des autres et ne forment donc pas un syncytium fonctionnel. Cependant ces muscles peuvent produire des mouvements fins contrairement aux muscles viscéraux.
Ultrastructure
Le cytoplasme des cellules musculaires lisses présente une zone bien définie qui contient les organites de la cellule (coiffant les deux pôles du noyau) et une autre qui occupe la plus grande partie de la cellule et qui contient les myofilaments. Les myofilaments d’actine (myofilaments fins), visibles en microscopie électronique, sont groupées en faisceaux irréguliers orientés selon le grand axe de la fibre. Ils sont associés à des molécules de tropomyosine et sont dépourvus de troponine. Les myofilaments épais de myosine ne sont pas visibles en microscopie électronique et leur mise en évidence nécessite des techniques de marquage particulières. Les protéines contractiles (myofilaments de myosine et d’actine) sont attachées à des zones denses constituées d’alpha-actinine qui sont soit dispersées dans le cytoplasme soit accolées à la face interne de la membrane plasmique. Des filaments intermédiaires constitués de desmine et de vimentine sont également attachés à ces zones denses.
Couplage excitation-contraction
Les phénomènes moléculaires de la contraction des fibres musculaires lisses nécessitent la présence de calcium. Un afflux de calcium sous sa forme ionique (Ca2+) provenant soit du réticulum endoplasmique soit de l’espace extra-cellulaire via les canaux calciques voltage et/ou ligand dépendants du domaine calvéolaire de la membrane plasmique. Le domaine calvéolaire est la portion de membrane plasmique qui présente de petites invaginations, les cavéoles ou vésicules plasmalemmales. Le calcium qui afflue dans la fibre musculaire lisse se lie à la calmoduline, une protéine de liaison du calcium (calcium-binding protein). Le complexe calcium-calmoduline qui s’est formé active une enzyme, la kinase des chaînes légères de myosine. Cette kinase permet la phosphorylation d’une des deux chaînes de myosine légères de chaque tête de myosine par l’utilisation de l’ATP. Cette phosphorylation permet de démasquer le site de liaison de l’actine sur la tête de myosine lourde. La liaison de l’actine avec la myosine induit la contraction de la fibre musculaire lisse.
Muscle strié cardiaque
Description macro et microscopique
Le muscle strié cardiaque est un muscle creux constitué de myocytes de contraction involontaire, rythmique et automatique qui forment un réseau tridimensionnel dans le myocarde. Les cardiomyocytes ont une forme de cylindre dont les extrémités forment des bifurcations avec les cellules adjacentes formant ce réseau tridimensionnel. Ils possèdent chacun un noyau central allongé dans le sens du grand axe de la cellule. Les striations observées dans le sarcoplasme des cardiomyocytes sont semblables à celles observées dans le muscle strié squelettique. Les extrémités des fibres adjacentes sont accolées l’une à l’autre au niveau d’une structure appelée disque intercalaire.
Métabolisme et vascularisation
Le muscle cardiaque est très dépendant du métabolisme oxydatif et est continuellement actif. Par conséquent, le flux sanguin nécessaire au fonctionnement de celui –ci est important. Ainsi de nombreux capillaires sont observés sur une coupe de muscle cardiaque.
Innervation
L’innervation du muscle cardiaque est de deux types : l’innervation intrinsèque et l’innervation extrinsèque.
La première, effectuée par l’intermédiaire du tissu nodal, est essentielle au contrôle et à la régulation de la contraction du myocarde et est responsable de l’automatisme cardiaque. Ce tissu est constitué de différents types cellulaires en fonction de leur topographie : les cellules nodales sont réparties en nœuds (sino-auriculaire et atrio-ventriculaire), en faisceaux (faisceau de His) et en réseau sous-encardique (réseau de Purkinje). L’influx qui permet de produire une contraction en rythme des fibres musculaires cardiaques naît au niveau du nœud sino-auriculaire, et modulé dans le nœud auriculo-ventriculaire et est transmis via le faisceau de His et le réseau de Purkinje.
L’innervation extrinsèque, quant à elle, intervient dans la régulation de l’activité du tissu nodal et ainsi peut moduler le rythme cardiaque. Cette innervation passe par l’intervention du système nerveux autonome sur la modulation de l’influx nerveux au niveau des nœuds.
Contraction
Le muscle cardiaque est un muscle de contraction involontaire, les myocytes le constituant se contractent de façon rythmique et automatique, on peut donc parler de syncytium fonctionnel. Cependant le muscle cardiaque n’est pas un syncytium au sens propre. Une cellule cardiaque est une cellule qui ne possède qu’un noyau, le syncytium fonctionnel s’effectue par l’intermédiaire des jonctions à trous qui permet aux cellules de communiquer entre elles.
Ultrastructure
L’ultrastructure du muscle cardiaque montre des caractères qui lui sont propres. Une région dépourvue de matériel et contenant divers organites cytoplasmiques est présente autour du noyau. Les mitochondries sont plus nombreuses et les grains de glycogène sont plus abondants que dans les fibres musculaires striées squelettiques. Le tubule T est localisé au niveau de la strie Z plutôt qu’au niveau de la jonction bande A-bande I comme cela est le cas dans le muscle strié squelettique. Au niveau des disques intercalaires, les membranes des cardiomyocytes adjacents sont parallèles l’une à l’autre au travers d’une importante série de plis. Cette structure permet de maintenir une importante cohésion de cellule à cellule. Ces disques intercalaires surviennent toujours sur des stries Z. Ainsi l’influence d’une unité contractile (le sarcomère) peut être transmise à l’unité suivante. Le long des flancs des fibres musculaires proche des disques intercalaires, les membranes des fibres adjacentes fusionnent sur de longues distances formant des jonctions à trous. Ces jonctions fournissent des ponts de faible résistance pour la propagation de l’excitation d’une fibre à l’autre. Elles permettent au muscle cardiaque de fonctionner comme un syncytium.
Nota Bene : Les termes tubule T, strie Z, bande A, bande I, sarcomère seront définis dans la partie descriptive du muscle strié squelettique.
Couplage excitation-contraction
La réponse contractile du muscle cardiaque commence peu après la dépolarisation de la membrane et dure environ 1.5 fois la durée du potentiel d’action. Il existe une période réfractaire absolue pendant laquelle une fibre musculaire cardiaque ne peut pas être à nouveau excitée, cette période s’étend de la dépolarisation de la membrane à la fin de la repolarisation c’est à dire environ 200 ms après la dépolarisation.
Le rôle du calcium (sous sa forme ionique) dans le couplage excitation-contraction est semblable à celui du muscle strié squelettique. Cependant, l’influx de calcium extracellulaire qui fait suite à l’activation des canaux dihydropyridine dans le système T est le précurseur des phénomènes moléculaires qui induisent la contraction
Muscle strié squelettique
Description macro et microscopique
Le muscle strié squelettique est par définition le muscle qui, par l’intermédiaire du tendon, se fixe au squelette et permet le mouvement de celui-ci dans une direction bien définie grâce à sa fonction essentielle de contraction. Il a été décrit, avec les premières observations au microscope optique sur du tissu en coupe longitudinale, comme étant un tissu présentant des striations à la fois transversales et longitudinales. Les fibres musculaires sont des cellules plurinucléées de forme polygonale dont les noyaux se situent en périphérie de la fibre, accolé à la membrane sarcoplasmique. Une fibre musculaire squelettique est un syncitium vrai c’est à dire un ensemble de protoplasmes ayant fusionnés entre eux. La fibre musculaire striée squelettique présente donc les caractéristiques fonctionnelles d’une cellule géante (1-5 cm de long, 10-100µm de diamètre).
Métabolisme et vascularisation
La vascularisation des fibres du muscle strié squelettique est effectuée comme tous les tissus via un réseau anastomotique de capillaires indispensable à l’oxygénation des tissus. Il s’agit d’un réseau à mailles rectangulaires autour des fibres musculaires. La densité de ce réseau est dépendante du type de la fibre concernée. Ce paragraphe sera développé dans un autre chapitre.
Innervation
Les fibres musculaires striées sont innervées par les motoneurones a dont le corps cellulaire est présent au niveau de la corne antérieure de la moelle épinière. Les unités motrices (ensemble formé par le neurone qui innerve la fibre, son axone et les fibres qu’il innerve) sont caractéristiques : les fibres innervées par un seul axone sont réparties aléatoirement dans le muscle.
Contraction
Le muscle strié squelettique est un muscle à contraction volontaire. Chacune des fibres le constituant est un syncytium structurel. L’intensité de la contraction du muscle dans son ensemble est fonction du nombre d’unités motrices qui ont été activées.
Ultrastructure
L’utilisation du microscope électronique pour définir l’ultrastructure du muscle strié squelettique a permis d’observer en détails les divers organites qui le compose, de décrire l’organisation des filaments contractiles et enfin d’en déduire la fonction de ces diverses structures dans la contraction de celui-ci.
Couplage excitation-contraction
Le couplage excitation-contraction est acétylcholine-dépendant. La dépolarisation de la membrane de la fibre musculaire striée squelettique déclenche la libération du calcium (stocké dans le réticulum sarcoplasmique, sous sa forme ionique) dans le sarcoplasme.
Un exemple de muscle strié à insertion conjonctive : le sphincter de l’urètre
Ces muscles sont constitués de fibres musculaires striées qui possèdent les mêmes caractéristiques que le muscle strié squelettique à quelques différences près.
Par exemple, le sphincter de l’urètre est constitué, autour de la lumière urétrale, d’une couche de muscle lisse semblable à la structure en double couche des tuniques musculaires tout au long de l’urètre et d’un anneau circulaire externe de muscle strié sur environ 70% de la longueur de l’urètre.
Le sphincter strié est constitué d’une couche circulaire constituée de quelques fibres musculaires séparées de la couche externe du sphincter par une bande de tissu conjonctif. Les fibres musculaires striées montrent un diamètre inférieur de l’ordre d’un facteur 3 à 5 à celui des fibres musculaires striées squelettiques. Les extrémités de ces fibres sont en contact direct avec le tissu conjonctif environnant ce qui différencie le muscle strié sphinctérien du muscle strié squelettique qui montre des jonctions myotendineuses. Cependant, le sphincter strié montre de grandes similitudes avec le muscle strié squelettique. Tout d’abord, la localisation cellulaire des protéines exprimées par le muscle strié squelettique montre une localisation semblable. L’innervation de ces fibres est acétylcholine-dépendant comme l’est l’innervation des fibres musculaires striées squelettiques. De plus l’aspect des jonctions neuromusculaires est très similaire à celui observé au niveau du muscle strié squelettique.
Néanmoins, les unités motrices montre une organisation différente de celle observée au niveau du muscle squelettique : les fibres musculaires innervées par un seul axone apparaissent contiguës. Le tissu conjonctif entourant les fibres musculaires striées délimite des faisceaux de fibres chevauchants agissant ensemble pour la fermeture de la lumière urétrale par l’intermédiaire du resserrement du muscle lisse.
